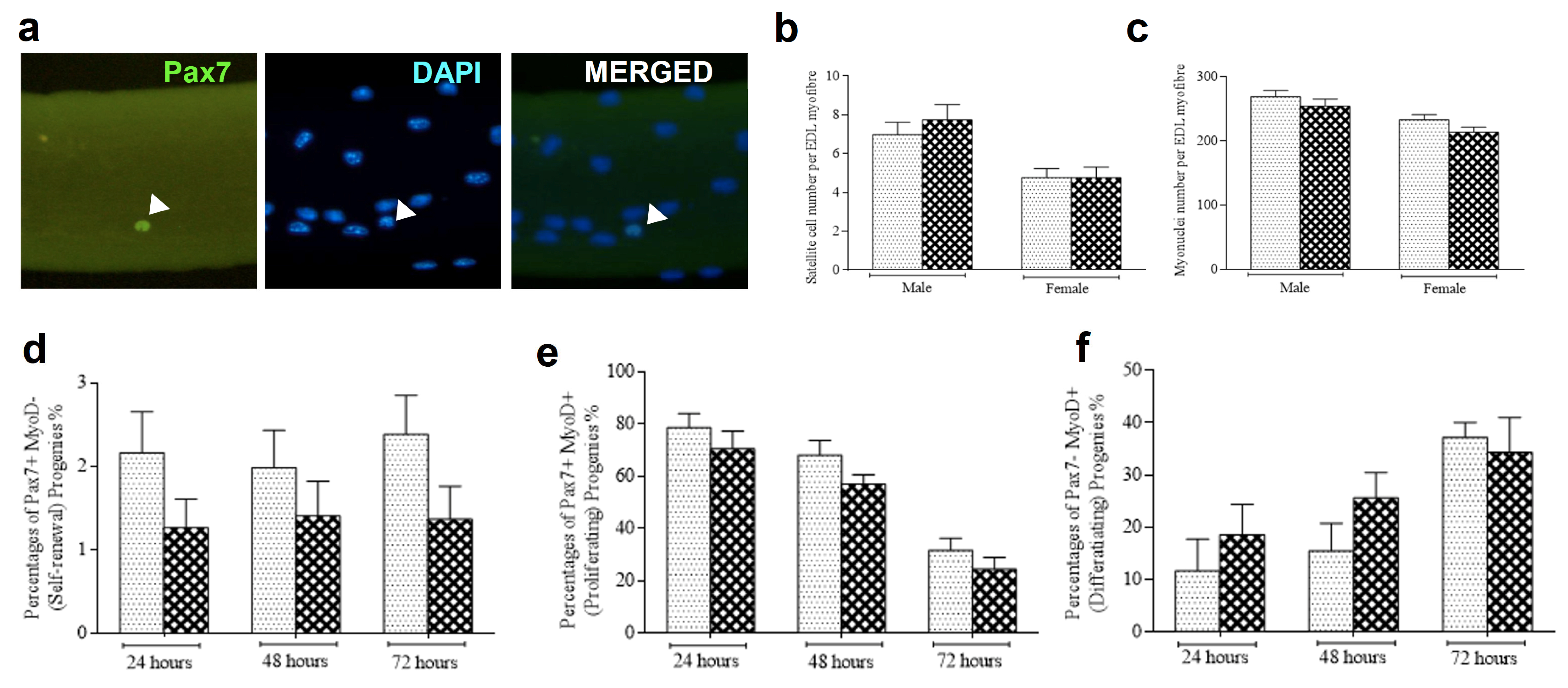
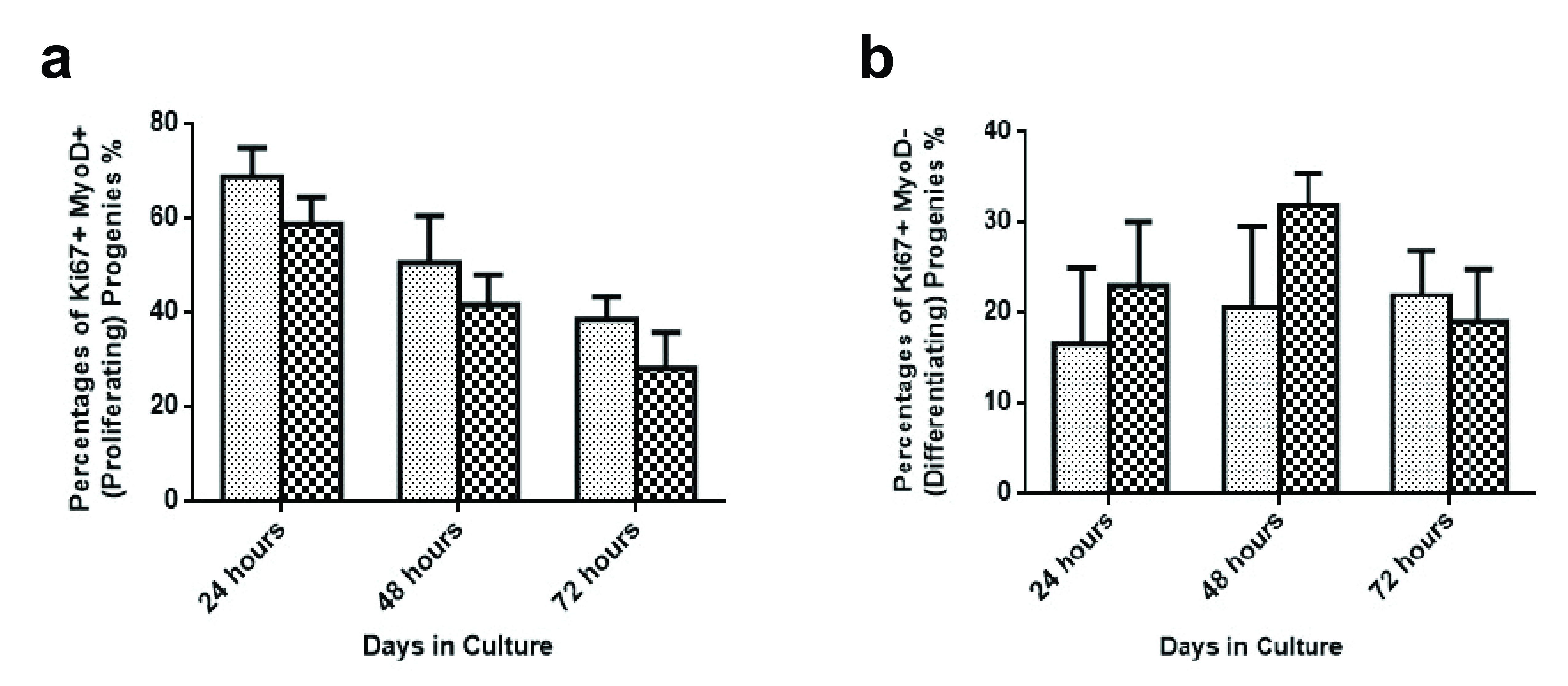
Acknowledgements: This work was
supported in part by funding from the Fundamental Research Grant Scheme
(04-01-15-1663FR) awarded to P-SC. C-LL was a recipient of the Malaysian
Ministry of Higher Education MyMaster scholarship. Monoclonal antibody, Pax7
was developed by A. Kawakami and obtained from the Developmental Studies
Hybridoma Bank developed under the auspices of the NICHD and maintained by the
Department of Biological Sciences, at the University of Iowa (Iowa City, IA,
USA).
Author
Contributions: All authors of this research paper have directly participated in the
planning, execution, or analysis of this study; CLL, K-HL, and P-SC designed
the experiments. C-LL, UB, and MP-YL performed experiments and analyzed the
data. IKSY, JS, and RR co-supervised experimentation. CLL, K-HL and P-SC
drafted the manuscript.
Conflicts
of Interest: The authors declare no conflict of interest.
References
1. Roper R.J., Reeves R.H. Understanding the basis for
Down syndrome phenotypes. PLoS Genet 2006, 2: e50. https://doi.org/10.1371/journal.pgen.0020050
2. National Center on Birth Defects and Developmental
Disabilities (NCBDDD) | CDC. National Center on Birth Defects and
Developmental Disabilities Home | NCBDDD | CDC. 2017. Available at: http://www.cdc.gov/ncbddd/birthdefects/downsyndrome.html. Accessed May 26, 2017.
3. Brault V., Duchon A.,
Romestaing C., Sahun I., Pothion S., Karout M., et al. Opposite phenotypes of muscle strength and locomotor
function in mouse models of partial trisomy and monosomy 21 for the proximal
Hspa13-App region. PLOS Genet 2015, 11: e1005062. https://doi.org/10.1371/journal.pgen.1005062
4. Lisi E.C., Cohn R.D. Genetic
evaluation of the pediatric patient with hypotonia: perspective from a
hypotonia specialty clinic and review of the literature. Dev Med Child Neurol 2011,
53: 586-599. https://doi.org/10.1111/j.1469-8749.2011.03918.x
5. Cowley P.M., Keslacy S.,
Middleton F.A., DeRuisseau L.R., Fernhall B., Kanaley J.A., DeRuisseau K.C.
Functional and biochemical characterization of soleus muscle in Down syndrome
mice: insight into the muscle dysfunction seen in the human condition. Am J Physiol Regul Integr Comp Physiol 2012, 303: R1251-R1260. https://doi.org/10.1152/ajpregu.00312.2012
6. Yin H., Price F., Rudnicki M.
Satellite cells and the muscle stem cell niche. Physiol Rev 2013, 93: 23-67.
https://doi.org/10.1152/physrev.00043.2011
7. Boldrin L., Morgan J.E. Human satellite cells:
identification on human muscle fibres. PLoS Curr 2013, 3: RRN1294. https://doi.org/10.1371/currents.RRN1294
8. Dumont N.A., Wang Y.X.,
Rudnicki M.A. Intrinsic and extrinsic mechanisms regulating satellite cell
function. Development 2015, 142: 1572-1581.
https://doi.org/10.1242/dev.114223
9. Tierney M.T., Sacco A. The
role of muscle stem cell-niche interactions during aging. Nat Med 2016, 22: 837-838.
https://doi.org/10.1038/nm.4159
10. Shefer G., Van de Mark D.P.,
Richardson J.B., Yablonka-Reuveni Z. Satellite-cell pool size does matter:
Defining the myogenic potency of aging skeletal muscle. Dev Biol 2006, 294: 50-66.
https://doi.org/10.1016/j.ydbio.2006.02.022
11. Shefer G., Rauner G., Yablonka-Reuveni
Z., Benayahu D. Reduced satellite cell numbers and myogenic capacity in aging
can be alleviated by endurance exercise. PLoS
ONE 2010, 5: e13307. https://doi.org/10.1371/journal.pone.0013307
12. Sago H., Carlson E., Smith D.,
Kilbridge J., Rubin E., Mobley W.C., Epstein C.J., Huang T.T. Ts1Cje, a partial trisomy 16 mouse model
for Down syndrome, exhibits learning and behavioral abnormalities. Proc Natl Acad Sci USA 1998, 95: 6256-6261. https://www.ncbi.nlm.nih.gov/pubmed/9600952
13. Liu B., Filippi S., Roy A.,
Roberts I. Stem and progenitor cell dysfunction in human trisomies. EMBO Rep 2015, 16: 44-62. https://doi.org/10.15252/embr.201439583
14.
Shimohata A., Ishihara K., Hattori S., Miyamoto H., Morishita H., Ornthanalai G., Raveau M., Ebrahim A.S., Amano K., Yamada K., Sago H., Akiba S., Mataga N., Murphy
N.P., Miyakawa T., Yamakawa K. Ts1Cje Down syndrome model mice exhibit
environmental stimuli-triggered locomotor hyperactivity and sociability
concurrent with increased flux through central dopamine and serotonin
metabolism. Exp Neurol. 2017,
293: 1-12. https://doi.org/10.1016/j.expneurol.2017.03.009
15. Ling K.H., Hewitt C.A., Tan
K.L., Cheah P.S., Vidyadaran S., Lai M.I., et
al. Functional transcriptome analysis of the postnatal brain of the Ts1Cje
mouse model for Down syndrome reveals global disruption of interferon-related
molecular networks. BMC Genomics 2014, 15: 624. https://doi.org/10.1186/1471-2164-15-624
16. Lim C.L., Ling K.H., Cheah
P.S. Isolation, cultivation and immunostaining of single myofibers- an improved
approach to study the behaviour of satellite cells. J Biol Methods 2018, 5:
e87. https://doi.org/10.14440/jbm.2018.219
17. Zammit P.S.,
Golding J.P., Nagata Y., Hudon V., Partridge T.A., Beauchamp J.R. Muscle
satellite cells adopt divergent fates. J Cell Biol 2004, 166:347-357.
https://doi.org/10.1083/jcb.200312007
18. Schlüter C.,
Duchrow M., Wohlenberg C., Becker M.H., Key G., Flad H.D., et al. The
cell proliferation-associated antigen of antibody Ki-67: a very large,
ubiquitous nuclear protein with numerous repeated elements, representing a new
kind of cell cycle-maintaining proteins. J Cell Biol 1993,
123: 513-522. https://doi.org/10.1083/jcb.123.3.513
19. Allen D.L., Roy R.R., Edgerton V.R. Myonuclear
domains in muscle adaptation and disease. Muscle
Nerve 1999, 22: 1350-1360. https://www.ncbi.nlm.nih.gov/pubmed/10487900
20. Brack A.S. Evidence that satellite cell decrement
contributes to preferential decline in nuclear number from large fibres during
murine age-related muscle atrophy. J Cell
Sci 2005, 118: 4813-4821. https://doi.org/10.1242/jcs.02602
21. Van der Meer S.F., Jaspers R.T., Degens H. Is the
myonuclear domain size fixed? J
Musculoskelet Neuronal Interact 2011, 11: 286-297. https://www.ncbi.nlm.nih.gov/pubmed/22130137
22. Neal A., Boldrin L., Morgan J.E. The satellite cell
in male and female, developing and adult mouse muscle: distinct stem cells for
growth and regeneration. PLoS ONE 2012, 7: e37950. https://doi.org/10.1371/journal.pone.0037950
23. Koopman R., Ly C.H., Ryall J.G. A metabolic link to
skeletal muscle wasting and regeneration. Front
Physiol 2014, 5: 32. https://doi.org/10.3389/fphys.2014.00032
24. Jang Y.C., Sinha M., Cerletti M., Dall'Osso C.,
Wagers A.J. Skeletal muscle stem cells: effects of aging and metabolism on
muscle regenerative function. Cold Spring Harb Symp Quant Biol 2011,
76: 101-111. https://doi.org/10.1101/sqb.2011.76.010652