
Cholinergic
modulation of hippocampal long-term potentiation in chronic cerebral hypoperfused
rats
Nor Fasihah Azam
1, Ryan Andrew Stanyard 2, Noorul Hamizah Mat 1
and Zurina Hassan 1,2,*
1
Centre for Drug Research, Universiti Sains Malaysia, 11800 Minden, Penang,
Malaysia.
2
School of Life Sciences, Keele University, Newcastle-under-Lyme, Staffordshire,
United Kingdom.
*
Correspondence: zurina_hassan@usm.my; Tel.: +604-6532726; Fax.: +604-6568669
Received: 26 June 2018; Accepted: 23 August 2018; Published: 6 September
2018
Edited by: Norshariza Nordin (Universiti Putra Malaysia,
Malaysia)
Reviewed by: Enoch Perimal (Universiti Putra Malaysia,
Malaysia); Raghava Sriramaneni (Univ. of Wisconsin, USA)
DOI: https://doi.org/10.31117/neuroscirn.v1i1.15
Abstract: Vascular dementia (VaD) is one
of the most common types of dementia in Alzheimer's disease (AD). Two-vessel
occlusion (2VO), also known as permanent bilateral occlusion of the common
carotid arteries, induces chronic cerebral hypoperfusion (CCH) in rats,
resulting in neuronal loss and inflammation (particularly in the cortex and
hippocampus). The 2VO rat model has been widely used to represent VaD
conditions similar to those seen in humans. Synaptic plasticity or long-term
potentiation (LTP) is one of the most important neurochemical foundations in
learning and memory, deficits of which occur as a result of VaD. The aim of
this study is to evaluate the role of cholinergic transmission in LTP
impairment of CCH rat model. There is a significant impairment of LTP following
the induction of 2VO surgery (p < .05). Treatment with oxotremorine
and tacrine cause significant enhancement of LTP and potentiation levels (p <
.05). There are also significant effects of paired-pulse facilitations when
treated with cholinergic agonists and baseline synaptic transmission with
increasing stimulation intensity (p < .0001). AChE activity was only
found to increase significantly in the hippocampal region (p < .05).
The role of cholinergic neurotransmission has been clearly demonstrated in
LTP impairment of the CCH rat model. Augmentation of synaptic transmission was
clearly observed in this model via changes of basal synaptic transmission and
neurotransmitter release presynaptically.
Keywords: vascular dementia; long-term potentiation; tacrine;
oxotremorine; carbachol;
©2018 by Azam et al for use and distribution in accord with the Creative
Commons Attribution (CC BY-NC 4.0) license (https://creativecommons.org/licenses/by-nc/4.0/),
which permits unrestricted non-commercial use, distribution, and reproduction
in any medium, provided the original author and source are credited.
1. Introduction
Vascular dementia (VaD) is one of the most common
types of dementia in Alzheimer's disease (AD). In 2015, it was estimated that
46.8 million people worldwide are living with dementia, with numbers predicted
to double every 20 years [1]. VaD typically develops when the blood
supply to the brain is reduced or blocked by damage to the vascular system [2]. Continuous reduction of cerebral blood
flow (CBF) affects neural structures regulating memory and cognitive processes
and eventually leads to the development and progression of VaD [3]. However, there are currently no approved
drugs that can stop the progression of VaD. Even though the accurate mechanisms
involved in VaD warrant further study, evidence is accumulating in various
experimental models of cerebral ischemia, which indicate that cholinergic
dysfunction may be implicated in the pathogenesis of this condition [4,5]. Furthermore, choline acetyltransferase (CAT) was
found to be significantly reduced in the brain of post-mortem VaD patients in
all areas except for the thalamus [6]. In addition, acetylcholinesterase (AChE)
activity has been found to be significantly higher in VaD rats compared to SHAM-surgery
rats [7]. Hence, regulating the cholinergic
pathways may prove to be an effective therapeutic strategy in attenuating VaD.
Two-vessel occlusion (2VO), also known as permanent
bilateral occlusion of the common carotid arteries, induces chronic cerebral
hypoperfusion (CCH) in rats, resulting in neuronal loss and inflammation
particularly in the cortex and hippocampus [8,9] with associated impairments in learning and memory [3,10]. In addition, occlusion of the vessels in the rat
causes a dramatic fall in CBF, creating hypoxic-ischemic conditions. This
condition closely resembles the reduced CBF in human ageing and dementia [3]. Moreover, this rat model has been
considered a representational model of VaD relative to the symptomatology seen
in humans [11,12].
Synaptic plasticity is defined as the ability of the
synapses to strengthen or weaken over time in response to neuronal stimulation.
Synaptic plasticity is one of the most important neurochemical foundations in
learning and memory. Memory has been postulated as a representation of widely
interconnected synapses within the brain [13]. The most studied model of synaptic
plasticity is that of long-term potentiation (LTP). LTP has been widely
explored to examine the synaptic strength between two neurons both in vivo
and in vitro [13,14]. It has been suggested that LTP has roles in both
learning and memory functions [15,16]. Previous studies have shown that the induction of
CCH in the VaD rat model cause impairments in LTP [17,18]. Therefore, maintaining the strength of synaptic
plasticity via the cholinergic pathway may offer a potential therapeutic
strategy for the prevention of cognitive deficits in VaD.
Activation and restoration of cholinergic function
remains a major objective in the development of pharmacological approaches
towards the treatment of cognitive dysfunctions associated with ageing and
dementia [19,20]. Acetycholinesterase inhibitors (AChEI) such as
tacrine have been shown to ameliorate the cognitive impairments seen in AD
patients [21]. In addition, cholinergic agonists such as
oxotremorine have been shown to increase the steady state concentration of
brain ACh [22] and treat the spatial memory deficits in
rats [23]. Another cholinergic drug, carbachol, has
been seen to increase brain activity via the production of theta rhythms in the
rat brain [24]. However, the effect of these drugs on in
vivo LTP has not been extensively studied. Therefore, the present study
seeks to investigate the role of the cholinergic system in LTP in the VaD rat
model.
2. Materials and Methods
2.1 Animal and housing conditions
All experimental procedures and animals were subjected
to approval by the Animal Ethics Committee, USM, Penang, Malaysia with the reference
number USM/Animal Ethics Approval/2014 (91) (538). Male Sprague-Dawley rats
weighing between 250-300g were obtained from the Animal Research & Service
Center (ARASC), USM, Penang, Malaysia. The rats were kept in plastic cages,
with ventilated stainless-steel covers, and were given one week to adjust to
conditions (maintained at 24 ± 1℃ and a 12:12 light/dark cycle) before
experiments commenced. The rats were fed with standard rodent food pellets and
water which were allowed ad libitum.
2.2 Surgery of 2-vessel occlusion
The rats were divided into two equal groups, the SHAM-operated
group and 2VO group. Permanent occlusion of the rat's common carotid arteries
was performed as described previously [25]. The rats were anaesthetized with a
mixture of ketamine hydrochloride (80 mg/kg; Troy Laboratories Pty. Ltd.,
Australia) and xylazine hydrochloride (10 mg/kg; Troy Laboratories Pty. Ltd.,
Australia) intraperitoneally, and the body temperature was maintained at 37 °C
using a homeothermic blanket (Harvard Apparatus, USA). The common carotid
arteries (CCA) were exposed via a ventral midline incision and carefully
isolated from the surrounding connective tissue and vagus nerves. Next, both
common carotid arteries were permanently ligated with suture silk 6/0 (UNIK
Surgical Sutures, Taiwan). The skin incision was closed and the rat was kept in
a cage at room temperature (25 ℃). The SHAM-operated control rats underwent
the same procedure, but without ligation. All rats were kept individually in
their cages and were given one week to recover prior to commencement of LTP
experiments.
2.3 Electrophysiological recording
The rats were anaesthetized with urethane (Sigma; 2.0
g/kg, administered in four doses 0.5 g/kg 20 min apart; supplements of 0.5 g/kg
were given when necessary) intraperitoneally before being secured on the
stereotaxic frame. Local analgesic xylocaine (5mg/kg; AstraZeneca, Australia)
was administered subcutaneously on the head. The rats were placed on the
stereotaxic apparatus and the ear bar was fixed into the auditory canal of each
ear respectively. The rat's body temperature was maintained between 36 ℃ and
37 ℃ using a homeothermic blanket (Harvard Apparatus). Subsequently, a small
incision on the head was made, and the connective tissue adhering to the bone
was carefully removed. The bone surface was cleared by using 30% hydrogen
peroxide (Sigma) until bregma and lambda were clearly visible. Two small holes
were drilled on the skull overlying the hippocampal CA1 (AP: -4.2. ML: +3, V: -3)
and CA3 (AP: -4.2, ML: -3, V: -4) regions, and two small holes were drilled
overlying the frontal region for placement of ground and reference electrodes.
All stereotaxic placements were based on the anatomical rat brain atlas of
Paxinos and Watson [26].
After drilling, a recording electrode (Perfluoroalkoxy
[PFA] Insulated Steel Wire, A-M Systems, USA) was lowered ventrally into the
CA1 region and a concentric, bipolar stimulation electrode (SNE 100,
MicroProbes, USA) was used to stimulate the CA3 region. The final ventral
placement of the CA3 stimulating electrode and recording electrode were
adjusted to yield maximum field excitatory postsynaptic potential (fEPSP) amplitude
in stratum pyramidale of CA1 in response to CA3 stimulation. The CA3
stimulation electrode was connected to a stimulus isolator providing constant
current output (ML 180 Stimulus Isolator, ADInstruments, Australia). The
recording, ground and reference electrodes were connected to an amplifier
(Model 1800, A-M Systems Inc., Sequim, WA; half amplitude filters set at 0.1-500
Hz) which digitised at 100 Hz and stored the recorded signal for offline
analysis (PowerLab 16/s System, ADInstruments, Australia).
For each rat, the intensity required to elicit the
maximal fEPSP amplitude was determined via input-output curves. The stimulation
was given between 0.1 mA and 1.0 mA (in 0.1 mA increments). The stimulation
intensity eliciting 50-60% of maximal fEPSP amplitude was used for the
remainder of the experiment. Paired-pulse facilitation was measured by
delivering pairs of stimulation pulses at inter-stimulus intervals of 20, 50,
100, 200, 500 and 1000 ms.
For LTP experiments, a baseline of the fEPSP was recorded
over a 60-minute period. Baseline recordings were considered stable when fEPSP
amplitude stayed within a range of 90-100% of the average fEPSP amplitude over
the 60-minute baseline period. Once a stable baseline was established, theta
burst stimulation (TBS) was applied as a train of ten bursts (each burst
consisting of 5 pulses at 100 Hz), repeated every 200 ms for each burst.
Following the TBS, single pulse stimulation of the CA3 continued every 30 sec
for a 3-hour recording period.
2.4 Drugs administration
In order to assess the role of cholinergic
transmission in the 2VO rat model, separate groups of rats received either
oxotremorine (Oxo) 0.1 mg/kg [27], tacrine (Tac) 3 mg/kg [20], or carbachol (Cch) 10µg/kg [28]. All drugs were dissolved in saline and
injected intraperitoneally (i.p). Oxotremorine, tacrine and carbachol were
obtained from Sigma. The rats were divided into groups as followed: Group 1:
SHAM + saline; Group 2: 2VO + saline; Group 3: SHAM + Oxo; Group
4: 2VO + Oxo; Group 5: SHAM + Cch; Group 6: 2VO + Cch; Group
7: SHAM + Tac; Group 8: 2VO + Tac. Each group consists of 5 to 6
rats. The drugs were given intraperitoneally before the start of the
experiments.
2.5 Determination of cholinesterase in brain samples
Different groups of rats were used to determine the
level of cholinesterase enzyme in 2VO model. The cholinesterase activity was measured
in rat's brain samples on day 28 of 2VO surgery. The brain tissues were
extracted and homogenized. Protein concentration was determined using
Bradford's assay and was subsequently followed by a cholinesterase assay.
2.6 Tissue extraction
The rats were anaesthetized using carbon dioxide (CO2)
and decapitated. CO2 has been widely used for short-acting
anaesthesia and euthanasia for small laboratory animals [29]. The brains were removed immediately and dissected
according to the areas of interest. The areas selected were the hippocampus,
frontal cortex and remaining cortex, sliced using a using tissue block (Kopf
Instrument) on ice. Next, the brain tissue was homogenized (40% w/v) in ice
cold phosphate buffer (0.1M, pH 8.0) with glass/Teflon tissue homogenizer
(Thomas Scientific, USA). The tissues were centrifuged at 1000 g for 10 minutes
at 4 ℃ to obtain the supernatant [30].
2.7 Determination of cholinesterase activity
The assessment of cholinesterase activity was measured
using the method from Ellman and colleagues [31] with minor modifications [32]. Theoretically, the presence of esterase
in the brain samples would hydrolyse either acetylthiocholine or
butyrylthiocholine respectively to produce thiocholine. Next, 5-dithiobis-2-nitrobenzoate
(DTNB; 3mM) reacted with thiocholine produced from the hydrolysis of
acetylthiocholine/butyrylthiocholine, leading to the formation of a yellow
coloured product that could be read at 412nm using a spectrophotometer.
For determination of acetylcholinesterase (AChE)
activity, acetylthiocholine iodide (ACTi; 14mM) was used as a substrate,
whereas for determination of butyrylcholinesterase (BuChE), butyrylthiocholine
iodide (BCTi; 14mM) was used as a substrate. Specific butyrylcholinesterase
inhibitor i.e. iso-OMPA (tetraisopropyl pyrophosphoramide; 2mM) was used in
addition to acetylthiocholine. Brain samples were loaded into the wells and the
reaction was initiated by adding the substrate accordingly. The summary of
volume and components of this assay were described briefly (Table 1). The
microplate was shaken for ~5 seconds and incubated for 15 minutes at 37 ℃.
After incubation, the absorbance was read at 412 nm using a microplate reader
(Multiskan Go, USA). Both AChE and BuChE enzyme activity were expressed as net
absorbance to protein amount optical densities/mg [30]. Protein was determined using Bradford's
assay [33,34].
2.8 Data analysis
For input-output curve, the fEPSP amplitude was
computed by measuring the difference between activity immediately preceding
each stimulation pulse and the peak of the fEPSP elicited by the stimulation.
fEPSP amplitudes recorded during input-output curve experiments were analysed
with stimulus intensity as the within-subject's variable. Paired-pulse
facilitation was computed by measuring the ratio of the first and second fEPSP
in each pair (amplitude of second fEPSP/amplitude of first fEPSP). Ratios were
analysed with inter-stimulus intervals as the within-subjects variable.
Table 1: Summary of reaction volume and component for cholinesterase assay.
|
Test
|
Type of test
|
Volume of tissue (ml)
|
Volume of inhibitor (ml)
|
Volume of buffer/DTNBb (ml)
|
Volume and type of substrate
|
|
1
|
Substrate blanks
|
-
|
-
|
195
|
5 (ACTic)
|
|
2
|
Tissue blanks
|
10
|
-
|
190
|
-
|
|
3
|
BuChe
|
10
|
-
|
180
|
10 (BCTid)
|
|
4
|
AChe
|
10
|
10 (iso-OMPAa)
|
175
|
5 (ACTi)
|
a
iso-OMPA - 2mM solution; b DTNB - 3 mM solution; c ACTi -
14mM solution; d BCTi - 14mM solution.
For the LTP study, amplitude data was averaged over 10-min
epochs and normalised by dividing all data points by the average baseline fEPSP
amplitude (pre-TBS) of the respective animal. The data was expressed in terms
of the mean ± S.E.M. Maximal fEPSP amplitude was computed offline using
LabChart software (version 7.3.7, ADInstrument, Australia). In this study, the
fEPSP amplitude was used in reference to previous studies [35-37]. Statistical analyses were carried out using Prism 5
for Windows software version 5.01 (GraphPad Software Inc., USA). Comparison
between two groups was analysed using an unpaired t-test. A One-way ANOVA was
also used when comparing more than three groups. The comparison between 2
variables was done using a two-way ANOVA. Alpha was set at the 5% level,
therefore differences between conditions required p < 0.05 to
evidence statistical significance.
3. Results
3.1 Input-output curves
Figure 1A shows the mean fEPSP amplitudes for rats
administered with oxotremorine (0.1mg/kg), with fEPSP amplitude increasing
similarly in all groups with increasing stimulation intensity. There was a
significant group effect (F3, 340 =35.58, p < .0001) and
intensity effect (F9,340 = 81.98, p < .0001) but there was
no interaction effect (p > .05). Figure 2A shows the mean fEPSP
amplitudes of rats treated with carbachol (10µg/kg). Similarly, there was
significant group effect (F3,340 = 31.30, p < .0001) and
intensity (F9,340 = 91.12, p < .0001), however not the
interaction effects (p > .05). Treatment with tacrine (3mg/kg)
resulted in significant groups effect (F3,320 =34.98, p <
.0001) and intensity effects (F9,320 =69.03, p < .0001)
but not the interaction (p > .05; Figure 3A).
3.2 Paired-pulse facilitation
Two successive stimulation pulses were applied to CA3
at short stimulus intervals (20-1000 ms), the second of the two resulted in
fEPSPs in CA1 exhibiting a larger amplitude relative to the first fEPSP in each
pair (Figures 1C, 2C and 3C). This facilitation effect disappeared with longer
inter-stimulus intervals (200-1000ms). There was a significant main effect
between groups when treated with oxotremorine (F3,72 = 10.66, p
< .0001), carbachol (F3,72 = 12.44, p < .0001) and
tacrine (F3,60 = 6.74, p < .001). There are significant
effects of the main interaction when the rats were administered with
oxotremorine (F15,72 = 2.01, p < .05), or carbachol (F15,72
=1.88) but not with tacrine (p > .05). There was significant
effect of inter-stimulus interval in all groups (p < .05).
3.3 The role of cholinergic neurotransmission in LTP
of CCH rat model
Figure 4 depicts the changes of fEPSP amplitudes
following the administration of oxotremorine. The two ways ANOVA revealed a
significant improvement in LTP after the administration of oxotremorine
[Interaction: (F69,384=1.58, p < .005), between groups (F3,384=84.65,
p < .0001) and time effect (F23,384=34.33, p <
.0001)] (Figure 4A). In addition, the potentiation level of the 2VO rats
treated with oxotremorine significantly higher compared to 2VO alone (p <
.05; Figure 4C).
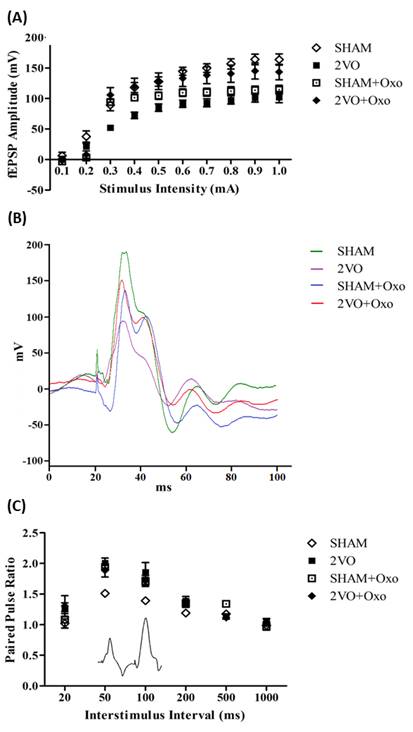
Figure 1. Field excitatory postsynaptic potentials (fEPSPs) in
CA1 elicited by CA3 stimulation in four experimental groups (2-vessel occlusion
[2VO], sham [SHAM], 2VO treated with oxotremorine [2VO+Oxo], sham treated with
oxotremorine [SHAM+Oxo]). (A) Input-output curves of all groups towards
increasing stimulation intensity. (B) The waveforms of fEPSP are illustrated
with the presence of stimulus artifact, fiber volley and fEPSP taken at 1 mA
stimulus intensity of each group. (C) Paired-pulse facilitation,
elicited by application of two successive stimulation pulses to CA3
(interstimulus intervals between 20 to 1000 ms), was measured as the amplitude
ratio of the second fEPSP divided by the first fEPSP in a pair. The insets
depict a typical fEPSP recorded in CA1 following a paired-pulse facilitation
applied to CA3 (A) as well as for an interstimulus interval of 50ms;
note the larger amplitude of the second fEPSP (B). Data expressed as
mean ± SEM of SHAM (n=5), 2VO (n=5), SHAM+Oxo (n=4) and 2VO+Oxo (n=4).
Another group of rats were tested with carbachol
(Cch), a cholinergic agonist. Administration of carbachol showed a significant
group effect (F3,360=30.46, p <.001) and time effect (F23,360=16.85,
p < .0001) however, no significant interaction effect was found (p
> .05; Figure 5A). Carbachol facilitated maintenance of LTP throughout the 3-hour
LTP recording in the 2VO group, but not in the SHAM group (p > .05).
However, no significant changes of the potentiation level were found between
groups at the end of the LTP recording (p > .05; Figure 5C).
In order to assess the contribution of cholinesterase
enzyme in the hippocampus, tacrine (Tac), a cholinesterase inhibitor was used. Administration of tacrine (3 mg/kg) caused a robust LTP induction following TBS
in both SHAM and 2VO groups (Figure 6A), with a significant interaction effect
(F69,336 = 3.68, p < .0001), groups effect (F3, 336 =
231, p < .0001) and time effect (F23,336 = 33.98, p
< .0001). Furthermore, both 2VO and SHAM treated with tacrine show
significant enhancement in potentiation level of fEPSP amplitudes compared to
non-treated groups (Figure 6C).
3.4 Determination of cholinesterase activity in CCH
rat model
Determination of the cholinesterase activity was done
on day 28 following the onset of the 2VO surgery. An unpaired Student t-test
was conducted to measure the activity between 2VO and SHAM groups at different
brain area i.e. hippocampus, frontal cortex and remaining cortex. The
level of AChE enzyme activity in 2VO group at the hippocampus region (p
< .05) was significantly higher compared to the other regions (Figure 7A).
In addition to AChE, level of BuChE enzyme activity was also measured in the
same regions. However, no significant changes were found in any of the
respective regions (p > .05; Figure 7B).

Figure 2. Field excitatory postsynaptic potentials (fEPSPs) in
CA1 elicited by CA3 stimulation in four experimental groups (2-vessel occlusion
[2VO], sham [SHAM], 2VO treated with carbachol [2VO+Cch], sham treated with
carbachol [SHAM+Cch]). (A) Input-output curves of all groups towards
increasing stimulation intensity. (B) The waveforms of fEPSP are
illustrated with the presence of stimulus artifact, fiber volley and fEPSP
taken at 1 mA stimulus intensity of each group. (C) Paired-pulse
facilitation, elicited by application of two successive stimulation pulses to
CA3 (interstimulus intervals between 20 to 1000 ms), was measured as the
amplitude ratio of the second fEPSP divided by the first fEPSP in a pair. Data
expressed as mean ± SEM of SHAM (n=5), 2VO (n=5), SHAM+Cch (n=4) and 2VO+Cch
(n=4).

Figure 3. Field excitatory postsynaptic potentials (fEPSPs) in
CA1 elicited by CA3 stimulation in four experimental groups (2-vessel occlusion
[2VO], sham [SHAM], 2VO treated with tacrine [2VO+Tac], sham treated with
tacrine [SHAM+Tac]). (A) Input-output curves of all groups towards
increasing stimulation intensity. (B) The waveforms of fEPSP are
illustrated with the presence of stimulus artifact, fiber volley and fEPSP
taken at 1 mA stimulus intensity of each group. (C) Paired-pulse
facilitation, elicited by application of two successive stimulation pulses to
CA3 (interstimulus intervals between 20 to 1000 ms), was measured as the amplitude
ratio of the second fEPSP divided by the first fEPSP in a pair. Data expressed
as mean ± SEM of SHAM (n=5), 2VO (n=5), SHAM+Tac (n=4) and 2VO+Tac (n=4).
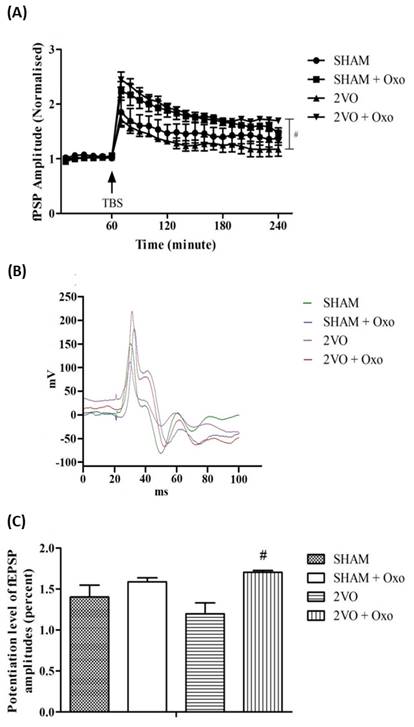
Figure 4. Changes of fEPSP amplitude following TBS in 2VO and
sham-operated group treated with oxotremorine (0.1mg/kg). The fEPSP amplitude
was plotted as a percent change against the baseline. (A) Each point
represents the changes of LTP following the TBS after the administration of
oxotremorine (0.1mg/kg). (B) The waveforms of fEPSP are illustrated
with the presence of stimulus artifact, fiber volley and fEPSP taken after 2
hours following TBS of all groups. (C) The potentiation level of LTP at
the end of the recordings which was taken during the last 60 minutes of the 3
hours LTP recording following the TBS. Data are expressed as mean ± S.E.M,* p
< 0.05 compared to sham-operated group and # p < 0.05 compared to 2VO
from 2VO (n=5), sham (n=5), 2VO + Oxo (n=4) and sham + Oxo (n=4) groups.
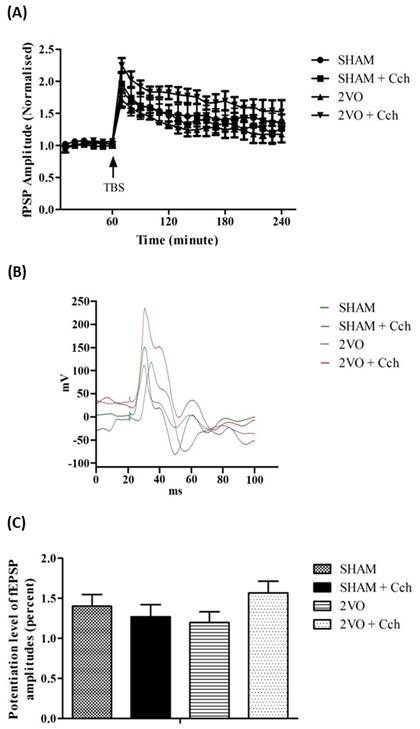
Figure 5. Changes of fEPSP amplitude following TBS in 2VO and
sham-operated group treated with carbachol (10 µg/kg). The fEPSP amplitude was
plotted as a percent change against the baseline. (A) Each point
represents the changes of LTP following the TBS after the administration of carbachol
(1 µg/100g). (B) The waveforms of fEPSP are illustrated with the
presence of stimulus artifact, fiber volley and fEPSP taken after 2 hours
following TBS of all groups. (C) The potentiation level of LTP at the
end of the recordings which was taken during the last 60 minutes of the 3 hours
LTP recording following the TBS. Data are expressed as mean ± S.E.M,* p <
0.05 compared to sham-operated group and # p < 0.05 compared to 2VO from 2VO
(n=5), sham (n=5), 2VO + Cch (n=4) and sham + Cch (n=4) groups.
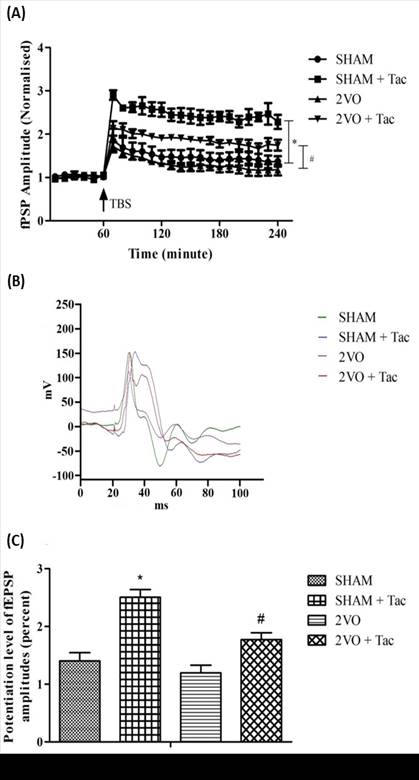
Figure 6. Changes of fEPSP amplitude following TBS in 2VO and
sham-operated group treated with tacrine (3 mg/kg). The fEPSP amplitude was
plotted as a percent change against the baseline. (A) Each point
represents the changes of LTP following the TBS after the administration of
tacrine (3 mg/kg). (B) The waveforms of fEPSP are illustrated with the
presence of stimulus artifact, fiber volley and fEPSP taken after 2 hours
following TBS of all groups. (C) The potentiation level of LTP at the
end of the recordings which was taken during the last 60 minutes of the 3 hours
LTP recording following the TBS. Data are expressed as mean ± S.E.M,* p <
0.05 compared to sham-operated group and # p < 0.05 compared to 2VO from 2VO
(n=5), sham (n=5), 2VO + Tac (n=4) and sham + Tac (n=4) groups.
4. Discussion
In this paper, we investigate the role of cholinergic
transmission in cellular and molecular mechanisms altering synaptic strength in
chronic cerebral hypoperfused rats. LTP is the most commonly studied at the
synapse between the Schaffer collateral axons of the CA3 neurons and the CA1
pyramidal cell [38]. In order to evaluate the basal synaptic
transmission at the CA1 region of the rat after stimulating the CA3 region of
the Schaffer-collateral pathway, an input-output curve was plotted. The fEPSP
amplitudes in input-output curve reveal significant effects of the experimental
and SHAM models treated with cholinergic drugs (Figures 1A, 2A and 3A). Many
aspects of the baseline synaptic transmission at CA3-CA1 synapses indicate
responses analogous to the SHAM group. Previous research studies indicate that
there was no impairment of basal synaptic transmission in the 2VO model [39,40]. On the contrary, others reported that induction of
2VO caused significant impairment of basal synaptic transmission [41]. The latter was found to be in line with
our findings. The reason for these discrepancies may be due to some alterations
in the molecular mechanisms involved during post-surgery, or indeed variation
between animals in terms of species or age [18].
Treatments with oxotremorine and carbachol showed
significant effects on the level of paired-pulse facilitation (PPF) as an
indirect measure of calcium levels and transmitter release from the presynaptic
terminal. The degree of PPF is inversely proportional to the release of
neurotransmitter from presynaptic terminals [42]. Therefore, altered levels of LTP
following the 2VO surgery are not likely to result from changes in presynaptic
release. Hence, we propose that a postsynaptic (rather than a presynaptic)
mechanism predominates following the 2VO surgery. However, the altered LTP
following cholinergic agonists (i.e oxotremorine and carbachol) may
derive from both pre- and postsynaptic mechanisms. On the contrary, treatment
with tacrine may involve the postsynaptic mechanism given that the drug depends
on the level of AChE enzymes present in the synaptic cleft.
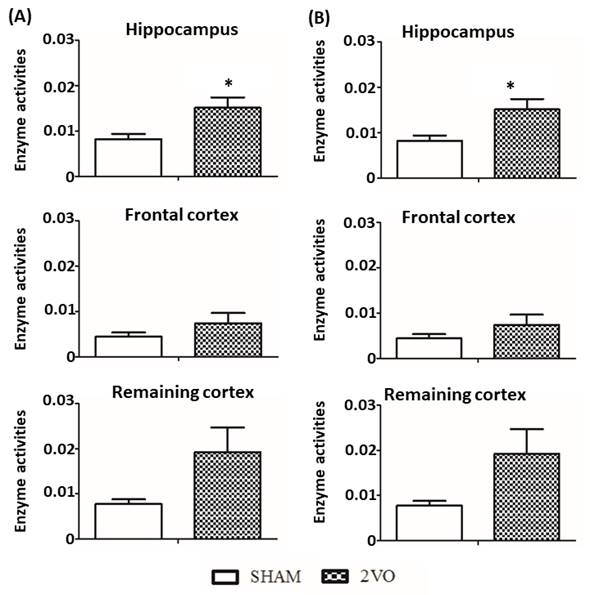
Figure 7. Determination of (A) AChE and (B) BuChE
activity between 2VO and sham-operated group on day 28 after the onset of the
surgery. Data are expressed as mean ± S.E.M with n=5 each group. The data was
analyzed using unpaired student t-test for each region.
As evidenced in Figure 4A, the LTP of the 2VO group
was significantly impaired compared to the SHAM-operated group following TBS.
The result is in accordance with the findings of previous studies suggesting
that 2VO caused significant reduction of LTP [9,18,40,43]. The causal mechanisms underlying LTP
impairment in this model are currently unclear. However, there are few possible
explanations that may contribute to the changes of the synaptic plasticity in
this model. Occlusion of both common carotid arteries will lead to reduction of
the cerebral blood flow and oxygen supplied to the brain. Consequently, this
would alter the biochemical metabolism of the neuron and potentially cause
neuronal cell death. This ischemic cell death may be prevalent in the
hippocampus, with respect to hippocampal pyramidal neurons. Further, permanent
impairment of LTP may develop in the rat hippocampal Schaffer-collateral CA1
which connects pyramidal neurons [44]. Furthermore, cAMP response element-binding
(CREB) protein was reported to be significantly reduced in the 2VO model [40,45,46].
CREB protein plays a role in neuronal plasticity and memory formation. In
addition, synaptophysin present in synaptic vesicles [17,47] and microtubule-associated protein 2 (MAP-2) [40,48] were significantly reduced in 2VO model.
Synaptophysin is a membrane protein in synaptic vesicles and localized at the
axon terminals while MAP-2 is a protein that functions for dendritic outgrowth,
branching and post-lesion dendritic remodelling [40]. These proteins are thought to play a
crucial role in maintaining the LTP and any malfunctioning of those proteins
may lead to long term depression (LTD) [49,50].
Induction of LTP is dependent on the protein
phosphorylation and dephosphorylation while maintaining LTP requires the
cascade of molecular process [14,49]. In the present study, induction of LTP was given by
theta-burst stimulation (TBS). TBS was given as 5-pulses burst at 100 Hz to mimic
complex-spike burst and repeated for 10 times at five bursts per second to
approximate the theta rhythm frequency. TBS is an effective stimulus for LTP
for several reasons. Firstly, it shows the pattern of neuronal firing (complex-spikes)
that occurred spontaneously during behaviour. Secondly, the optimal repetition
rate corresponded to hippocampal theta rhythm (indirectly correlating with
memory processes in EEG), the stimulation given will allow uncovering various
events that contribute to LTP induction [51]. Furthermore, TBS is highly sensitive
towards LTP. For example, TBS is vulnerable in aging [52], experimentally-induced stress [53], has become more sensitive to the effect
of brain-derived neurotrophic factor (BDNF) [54], serotonin [55] or adenosine receptor-dependent signalling
mechanisms [56]. Altogether, TBS of LTP may provide a
better model for the memory effects than tetanus-induced LTP.
Previous studies had shown that induction of LTP at
the Schaffer collateral-CA1 synapse was enhanced through activation of
cholinergic receptors [57-59]. For example, a study by Shimoshige and colleagues (1997)
had examined the effects of carbachol, acting as a muscarinic receptor agonist
with respect to LTP. They reported that carbachol significantly increased the
magnitude of LTP [58]. In addition, a study by Doralp and Leung
(2008) on the cholinergic modulation of hippocampal CA1 LTP resulted in
enhanced LTP in vivo when the rats were treated with the
anticholinesterase drug, serine sulfate [59]. Another study by Ovsepian and colleagues
(2004) examined the role of cholinergic on the induction of LTP using non-selective
and selective muscarinic antagonist drugs, scopolamine and pirenzepine
respectively [60]. These authors concluded that the CA1 area
is a muscarinic ACh receptor dependent and medial septum pre-conditioning
transiently facilitates LTP induction in these synapses. Furthermore,
muscarinic antagonist atropine caused significant decrease of LTP after the
strong tetanization [61]. In the present study, the non-selective
muscarinic agonist oxotremorine is able to enhance LTP in the CA1 of the 2VO
rat. The potentiation level of LTP at the end of the recordings in 2VO treated
with oxotremorine remained higher compared to 2VO group alone (Figure 4).
Carbachol, a cholinergic agonist and tacrine, an
anticholinesterase was also used in the present study. Both drugs enhanced fEPSP
amplitude in the CA1 region of the 2VO rat. However, the potentiation level of
carbachol was unchanged when in the sham group, suggesting that the cholinergic
receptors were not affected (Figure 5). The administration of tacrine produced
robust LTP in both 2VO and sham groups (Figure 6). Thus, there is contribution
of AChE in this model. The level of ACh in the brain (especially in
hippocampus) may be low due to rapid hydrolysis by AChE [62]. This could be seen via the AChE assay in the
hippocampal region (Figure 7A). It can therefore be suggested that an increased
level of AChE can lead to rapid hydrolysis of ACh to choline and acetate. Thus,
it may reduce the binding of ACh to the cholinoceptors. As a consequence,
critical decrease in the level of ACh may contribute to the cognitive deficits
evident in the 2VO rats [63]. As a summary, these results imply that
central cholinergic impairment induced by 2VO may play a critical role in the
development of cognitive dysfunction. Therefore, ameliorating central
cholinergic dysfunction may be an auspicious strategy for managing cognitive
impairment associated with cerebral hypoperfusion.
Impairment in learning and memory is one of the
characteristic features of dementia in AD and VaD. The Morris water maze has
become one of the most extensively accepted test models for testing learning
and memory in rodents [64]. Data from our laboratory have
demonstrated that 2VO rats showed significantly longer escape latencies during
the 5-days training period, as well as during probe trials [10]. This suggest that this model exhibits
spatial learning and memory deficits. Impairment in memory and learning was associated
with the decreased level of acetylcholine (ACh) [5] and also increase level of AChE [65] in the brain. ACh is a well-known
neurotransmitter that functions in many parts of human body and especially
involved in some central functions i.e. walking, cognition control or
memory [66]. Since the discovery of rapid colorimetric to
determine the AChE activity by Ellman and colleagues, there are growing number
of studies have been developed using this approach as a validation method for
cognitive functions [31]. In the present study, the cholinesterase
enzyme tested resulted in significant increase of AChE in hippocampus region
but not other region (Figure 7A). However, no significant changes were found
for BuChE (Figure 7B).
In summary, our results showed the contributions of
AChE inhibitor, tacrine and cholinergic agonists, oxotremorine and carbachol in
ameliorating the LTP impairment in CCH rat model. The CCH rat model may be
correlated with the increase of the cholinesterase enzyme activity. High level
of cholinesterase enzyme activity was found to be the major problem in memory
impairment of VaD model. Therefore, targeting the cholinergic pathways may be a
plausible manner of affecting the cognitive deficits in VaD. These findings are
useful for the development of new potential neuroprotective agents associated
with dementia or cerebral hypoperfusion targeting the cholinergic
neurotransmission.
5. Conclusions
The role of cholinergic neurotransmission has been
clearly demonstrated in LTP impairment of the VaD rat model. Drugs such as
tacrine, oxotremorine and carbachol are effective in improving the LTP deficit
in CCH model. Therefore, these results can be used as a guideline in producing
new therapeutic agents targeting on the cholinergic pathways.
Acknowledgements: This work was supported by the Malaysian Ministry of
Higher Education (MOHE), the Universiti Sains Malaysia special funding for the
Fundamental Neuroscience-Neurobehavior project (304/CDADAH/652201 /K134), the
Research University Individual (RUI) Grant (1001/ CDADAH/812177) and the USM
Fellowship Scheme for the postgraduate funding.
Author Contributions: NFA performed all the experiments in this study and
contributed to the writing of the manuscript. ZH developed the methods for LTP
and AChE assay experiments. ZH and RAS contributed to the research discussions.
NFA wrote the manuscript, which was revised by RAS, ZH and NHM. All authors
read and approved the final manuscript.
Conflicts of Interest: The authors declare no conflict of interest.
References
1.
Prince M, Wimo A,
Guerchet M, Ali G-C, Wu Y-T, Prina M. World Alzheimer Report 2015: The Global
Impact of Dementia - An analysis of prevalence, incidence, cost and trends.
2015. https://www.alz.co.uk/research/worldalzheimerreport2015summary.pdf
2.
Roman GC, T E, A W, L.
P, Chui HC. Subcortical ischaemic vascular dementia. The Lancet. 2002;
1(7): 426-436. https://www.ncbi.nlm.nih.gov/pubmed/12849365
3.
Farkas E, Luiten PGM,
Bari F. Permanent, bilateral common carotid artery occlusion in the rat: a
model for chronic cerebral hypoperfusion-related neurodegenerative diseases. Brain
Res Rev. 2007;54(1):162-180. https://doi.org/10.1016/j.brainresrev.2007.01.003
4.
Ma H, Mochida S. A
cholinergic model synapse to elucidate protein function at presynaptic
terminals. Neurosci Res. 2007; 57(4):491-498. https://doi.org/10.1016/j.neures.2006.12.015
5.
Ni JW, Matsumoto K, Li
HB, Murakami Y, Watanabe H. Neuronal damage and decrease of central
acetylcholine level following permanent occlusion of bilateral common carotid
arteries in rat. Brain Res. 1995;673(2):290-296. https://doi.org/10.1016/0006-8993(94)01436-L
6.
Gottfries CG, Blennow
K, Karlsson I, Wallin A. The Neurochemistry of Vascular Dementia. Dementia.
1994;5(3-4):163-167. http://www.ncbi.nlm.nih.gov/pubmed/8087172
7.
Kwon KJ, Kim MK, Lee
EJ, Kim JN, Choi B-R, Kim SY, et al. Effects of donepezil, an
acetylcholinesterase inhibitor, on neurogenesis in a rat model of vascular
dementia. J Neurol Sci. 2014;347(1-2):66-77. https://doi.org/10.1016/j.jns.2014.09.021
8.
Pereira FM, Ferreira
EDF, de Oliveira RMW, Milani H. Time-course of neurodegeneration and memory
impairment following the 4-vessel occlusion/internal carotid artery model of
chronic cerebral hypoperfusion in middle-aged rats. Behav Brain Res.
2012;229(2):340-348. https://doi.org/10.1016/j.bbr.2012.01.023
9.
Kocsis K, Knapp L,
Gellért L, Oláh G, Kis Z, Takakuwa H, et al. Acetyl-l-carnitine normalizes the
impaired long-term potentiation and spine density in a rat model of global
ischemia. Neuroscience. 2014;269:265-272. https://doi.org/10.1016/j.neuroscience.2014.03.055
10. Damodaran T, Hassan Z, Navaratnam V,
Muzaimi M, Ng G, Müller CP, et al. Time course of motor and cognitive functions
after chronic cerebral ischemia in rats. Behav Brain Res. 2014;275:252-258.
https://doi.org/10.1016/j.bbr.2014.09.014
11. Kitamura A, Fujita Y, Oishi N, Kalaria RN,
Washida K, Maki T, et al. Selective white matter abnormalities in a novel rat
model of vascular dementia. Neurobiol Aging. 2012;33(5):1012.e25-1012.e35.
https://doi.org/10.1016/j.neurobiolaging.2011.10.033
12. Amenta F, Di Tullio MA, Tomassoni D. The
cholinergic approach for the treatment of vascular dementia: evidence from pre-clinical
and clinical studies. Clin Exp Hypertens. 2002;24(7-8):697-713. https://www.ncbi.nlm.nih.gov/pubmed/12450245
13.
Larkman AU, Jack JJ.
Synaptic plasticity: hippocampal LTP. Curr Opin Neurobiol. 1995;5(3):324-334.
https://doi.org/10.1016/0959-4388(95)80045-X
14. Neves G, Cooke SF, Bliss T V. Synaptic
plasticity, memory and the hippocampus: a neural network approach to causality.
Nat Rev Neurosci. 2008;9(1):65-75. https://doi.org/10.1038/nrn3396
15. Xu B, Li XX, He GR, Hu JJ, Mu X, Tian S, et
al. Luteolin promotes long-term potentiation and improves cognitive functions
in chronic cerebral hypoperfused rats. Eur J Pharmacol . 2010;627(1-3):99-105.
https://doi.org/10.1016/j.ejphar.2009.10.038
16. Xu J, Wang Y, Li N, Xu L, Yang H, Yang Z. L-3-n-butylphthalide
improves cognitive deficits in rats with chronic cerebral ischemia. Neuropharmacology.
2012;62(7):2424-2429. https://doi.org/10.1016/j.neuropharm.2012.02.014
17. Wang Z, Fan J, Wang J, Li Y, Duan D, Du G,
et al. Chronic cerebral hypoperfusion induces long-lasting cognitive deficits
accompanied by long-term hippocampal silent synapses increase in rats. Behav
Brain Res. 2016;301:243-252. https://doi.org/10.1016/j.bbr.2015.12.047
18. Luo P, Lu Y, Li C, Zhou M, Chen C, Lu Q, et
al. Long-lasting spatial learning and memory impairments caused by chronic
cerebral hypoperfusion associate with a dynamic change of HCN1/HCN2 expression
in hippocampal CA1 region. Neurobiol Learn Mem. 2015;123:72-83. https://doi.org/10.1016/j.nlm.2015.05.005
19. Knopman DS. Cerebrovascular disease and
dementia. Br J Radiol. 2007;80(2):S121-127. https://doi.org/10.1259/bjr/75681080
20. Diez-Ariza M, Redondo C, García-Alloza M,
Lasheras B, Del Río J, Ramírez MJ. Flumazenil and tacrine increase the
effectiveness of ondansetron on scopolamine-induced impairment of spatial
learning in rats. Psychopharmacology (Berl). 2003;169(1):35-41. https://doi.org/10.1007/s00213-003-1467-1
21. Terry A V, Callahan PM, Hall B, Webster SJ.
Alzheimer's disease and age-related memory decline (preclinical). Pharmacol
Biochem Behav. 2011;99(2):190-210. https://doi.org/10.1016/j.pbb.2011.02.002
22. Bertrand N, Beley A. Effect of
oxotremorine, physostigmine, and scopolamine on brain acetylcholine synthesis:
A study using HPLC. Neurochem Res. 1990;15(11):1097-1100. https://doi.org/10.1007/BF01101710
23. Murakami Y, Ikenoya M, Matsumoto K, Li H,
Watanabe H. Ameliorative effect of tacrine on spatial memory deficit in chronic
two-vessel occluded rats is reversible and mediated by muscarinic M1 receptor
stimulation. Behav Brain Res. 2000;109(1):83-90. https://doi.org/10.1016/S0166-4328(99)00162-X
24. Natsume K, Kometani K. Desynchronization of
carbachol-induced theta-like activities by alpha-adrenergic agents in guinea
pig hippocampal slices. Neurosci Res. 1999;33(3):179-186. https://doi.org/10.1016/S0168-0102(99)00007-3
25. Castaño Guerrero Y, González Fraguela ME,
Fernández Verdecia I, Horruitiner Gutiérrez I, Piedras Carpio S. Changes in
oxidative metabolism and memory and learning in a cerebral hypoperfusion model
in rats. Neurologia. 2013;28(1):1-8. https://doi.org/10.1016/j.nrleng.2012.01.001
26. Paxinos G, Watson C. The Rat Brain in
Stereotaxic Coordinates, 6th Edition. 2006.
27. Almaguer-Melian W, Rojas-Reyes Y, Alvare A,
Rosillo JC, Frey JU, Bergado J A. Long-term potentiation in the dentate gyrus
in freely moving rats is reinforced by intraventricular application of
norepinephrine, but not oxotremorine. Neurobiol Learn Mem.2005;83(1):72-78.
https://doi.org/10.1016/j.nlm.2004.08.002
28. Wang M, Chen W-H, Zhu D-M, She J-Q, Ruan D-Y.
Effects of carbachol on lead-induced impairment of the long-term
potentiation/depotentiation in rat dentate gyrus in vivo. Food Chem Toxicol.
2007;45:412-418. https://doi.org/10.1016/j.fct.2006.08.025
29. Kohler I, Meier R, Busato A, Neiger-Aeschbacher
G, Schatzmann U. Is carbon dioxide (CO2) a useful short acting anaesthetic for
small laboratory animals? Lab Anim. 1999;33(2):155-161. https://doi.org/10.1258/002367799780578390
30. Mohr F, Zimmermann M, Klein J. Mice
heterozygous for AChE are more sensitive to AChE inhibitors but do not respond
to BuChE inhibition. Neuropharmacology. 2013;67:37-45. https://doi.org/10.1016/j.neuropharm.2012.11.001
31. Ellman GL, Courtney KD, Andres V,
Featherstone RM. A new and rapid colorimetric determination of
acetylcholinesterase activity. Biochem Pharmacol. 1961;7(2):88-95. https://doi.org/10.1016/0006-2952(61)90145-9
32. Padilla S, Lassiter TL, Hunter D.
Biochemical measurement of cholinesterase activity. Methods Mol Med.
1999; 22:237-245. https://doi.org/10.1385/0-89603-612-X:237
33. Kruger NJ. The bradford method for protein
quantitation. In: Walker J.M. (eds) Basic Protein and Peptide Protocols.
Methods in Molecular Biology™. Humana Press. 1994;32. https://doi.org/10.1385/0-89603-268-X:9
34. Bradford MM. A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing the principle
of protein-dye binding. Anal Biochem. 1976;72(1-2):248-254. https://doi.org/10.1016/0003-2697(76)90527-3
35. Dringenberg HC, Hamze B, Wilson A,
Speechley W, Kuo M-C. Heterosynaptic facilitation of in vivo thalamocortical
long-term potentiation in the adult rat visual cortex by acetylcholine. Cereb
Cortex. 2007;17(4):839-848. https://doi.org/10.1093/cercor/bhk038
36. Hager AM, Gagolewicz PJ, Rodier S, Kuo MC,
Dumont TC, Dringenberg HC. Metaplastic up-regulation of LTP in the rat visual
cortex by monocular visual training: Requirement of task mastery, hemispheric
specificity, and NMDA-GluN2B involvement. Neuroscience. 2015;293:171-186.
http://dx.doi.org/10.1016/j.neuroscience.2015.02.027
37. Hogsden JL, Rosen LG, Dringenberg HC.
Pharmacological and deprivation-induced reinstatement of juvenile-like long-term
potentiation in the primary auditory cortex of adult rats. Neuroscience.
2011;186:208-219. https://doi.org/10.1016/j.neuroscience.2011.04.002
38. Bliss T V, Collingridge GL. A synaptic
model of memory: long-term potentiation in the hippocampus. Nature. 1993;361(6407):31-39.
https://doi.org/10.1038/361031a0
39. Li Z, Zhou R, Cui S, Xie G, Cai W, Sokabe
M, et al. Dehydroepiandrosterone sulfate prevents ischemia-induced impairment
of long-term potentiation in rat hippocampal CA1 by up-regulating tyrosine
phosphorylation of NMDA receptor. Neuropharmacology. 2006;51(5):958-966.
https://doi.org/10.1016/j.neuropharm.2006.06.007
40. Zhu H, Zhang J, Sun H, Zhang L, Liu H, Zeng
X, et al. An enriched environment reverses the synaptic plasticity deficit
induced by chronic cerebral hypoperfusion. Neurosci Lett. 2011;502(2):71-75.
https://doi.org/10.1016/j.neulet.2011.04.015
41. Nagy D, Kocsis K, Fuzik J, Marosi M, Kis Z,
Teichberg VI, et al. Kainate postconditioning restores LTP in ischemic
hippocampal CA1: onset-dependent second pathophysiological stress. Neuropharmacology.
2011;61(5-6):1026-1032. https://doi.org/10.1016/j.neuropharm.2011.07.005
42. Schulz PE, Cook EP, Johnston D. Using
paired-pulse facilitation to probe the mechanisms for long-term potentiation
(LTP). J Physiol Paris. 1995;89(1):3-9. https://doi.org/10.1016/0928-4257(96)80546-8
43. Xu X, Li Z, Yang Z, Zhang T. Decrease of
synaptic plasticity associated with alteration of information flow in a rat
model of vascular dementia. Neuroscience. 2012;206:136-143. https://doi.org/10.1016/j.neuroscience.2011.12.050
44.
Otori T, Katsumata T,
Muramatsu H, Kashiwagi F, Katayama Y, Terashi A. Long-term measurement of
cerebral blood flow and metabolism in a rat chronic hypoperfusion model. Clin
Exp Pharmacol Physiol. 2003;30(4):266-272. https://doi.org/10.1046/j.1440-1681.2003.03825.x
45. Okuyama S, Shimada N, Kaji M, Morita M,
Miyoshi K, Minami S, et al. Heptamethoxyflavone, a citrus flavonoid, enhances
brain-derived neurotrophic factor production and neurogenesis in the
hippocampus following cerebral global ischemia in mice. Neurosci Lett.
2012;528(2):190-195. https://doi.org/10.1016/j.neulet.2012.08.079
46. Li H, Wang J, Wang P, Rao Y, Chen L.
Resveratrol Reverses the Synaptic Plasticity Deficits in a Chronic Cerebral
Hypoperfusion Rat Model. J Stroke Cerebrovasc Dis. 2015;25(1):122-128. https://doi.org/10.1016/j.jstrokecerebrovasdis.2015.09.004
47. Lin Q, Hai J, Yao L-Y, Lu Y.
Neuroprotective effects of NSTyr on cognitive function and neuronal plasticity
in rats of chronic cerebral hypoperfusion. Brain Res. 2010;1325:183-190.
https://doi.org/10.1016/j.brainres.2010.02.037
48. Institoris A, Farkas E, Berczi S, Sule Z,
Bari F. Effects of cyclooxygenase (COX) inhibition on memory impairment and
hippocampal damage in the early period of cerebral hypoperfusion in rats. Eur
J Pharmacol. 2007;574(1):29-38. https://doi.org/10.1016/j.ejphar.2007.07.019
49. Raymond CR. LTP forms 1, 2 and 3: different
mechanisms for the “long” in long-term potentiation. Trends Neurosci.
2007;30(4):167-175. https://doi.org/10.1016/j.tins.2007.01.007
50. Lisman J, Raghavachari S. Biochemical
principles underlying the stable maintenance of LTP by the CaMKII/NMDAR
complex. Brain Res. 2014;1621:51-61. https://doi.org/10.1016/j.brainres.2014.12.010
51. Larson J, Munkácsy E. Theta-burst LTP. Brain
Res. 2015;1621:38-50. https://doi.org/10.1016/j.brainres.2014.10.034
52. Bartsch T, Wulff P. The hippocampus in
aging and disease: From plasticity to vulnerability. Neuroscience.
2015;309:1-16. https://doi.org/10.1016/j.neuroscience.2015.07.084
53. Maroun M, Richter-Levin G. Exposure to
acute stress blocks the induction of long-term potentiation of the amygdala-prefrontal
cortex pathway in vivo. J Neurosci. 2003;23(11):4406-4409. https://doi.org/10.1523/JNEUROSCI.23-11-04406.2003
54. Lu Y, Christian K, Lu B. BDNF: a key
regulator for protein synthesis-dependent LTP and long-term memory? Neurobiol
Learn Mem. 2008;89(3):312-323. https://doi.org/10.1016/j.nlm.2007.08.018
55. Stäubli U, Xu FB. Effects of 5-HT3 receptor
antagonism on hippocampal theta rhythm, memory, and LTP induction in the freely
moving rat. J Neurosci. 1995;15(3 Pt 2):2445-2452. https://doi.org/10.1523/JNEUROSCI.15-03-02445.1995
56. Shahraki A, Stone TW. Blockade of
presynaptic adenosine A1 receptor responses by nitric oxide and superoxide in
rat hippocampus. Eur J Neurosci. 2004;20(3):719-728. https://doi.org/10.1111/j.1460-9568.2004.03502.x
57. Tanaka K, Ogawa N, Asanuma M, Kondo Y,
Nomura M. Relationship between cholinergic dysfunction and discrimination
learning disabilities in Wistar rats following chronic cerebral hypoperfusion. Brain
Res. 1996;729(1):55-65. https://doi.org/10.1016/0006-8993(96)00400-3
58. Shimoshige Y, Maeda T, Kaneko S, Akaike A,
Satoh M. Involvement of M2 receptor in an enhancement of long-term potentiation
by carbachol in Schaffer collateral-CA1 synapses of hippocampal slices. Neurosci
Res. 1997;27(2):175-180. https://doi.org/10.1016/S0168-0102(96)01147-9
59. Doralp S, Leung LS. Cholinergic modulation
of hippocampal CA1 basal-dendritic long-term potentiation. Neurobiol Learn
Mem. 2008;90(2):382-388. https://doi.org/10.1016/j.nlm.2008.05.013
60. Ovsepian S V., Anwyl R, Rowan MJ.
Endogenous acetylcholine lowers the threshold for long-term potentiation
induction in the CA1 area through muscarinic receptor activation: In vivo
study. Eur J Neurosci. 2004;20(5):1267-1275. https://doi.org/10.1111/j.1460-9568.2004.03582.x
61. Sokolov M V, Kleschevnikov AM. Atropine
suppresses associative LTP in the CA1 region of rat hippocampal slices. Brain
Res. 1995;672(1-2):281-284. https://doi.org/10.1016/0006-8993(94)01376-S
62.
Watanabe T, Takasaki K, Yamagata N, Fujiwara M, Iwasaki K. Facilitation
of memory impairment and cholinergic disturbance in a mouse model of
Alzheimer's disease by mild ischemic burden. Neurosci Lett.
2013;536:74-79. https://doi.org/10.1016/j.neulet.2012.12.041
63. Ray RS, Rai S, Katyal A. Cholinergic
receptor blockade by scopolamine and mecamylamine exacerbates global cerebral
ischemia induced memory dysfunction in C57BL/6J mice. Nitric Oxide.
2014;43:62-73. https://doi.org/10.1016/j.niox.2014.08.009
64. Farkas E, Institóris A, Domoki F, Mihály A,
Luiten PGM, Bari F. Diazoxide and dimethyl sulphoxide prevent cerebral
hypoperfusion-related learning dysfunction and brain damage after carotid
artery occlusion. Brain Res. 2004;1008(2):252-260. https://doi.org/10.1016/j.brainres.2004.02.037
65. Gupta S, Sharma B. Pharmacological
modulation of I1-imidazoline and a2-adrenoceptors in sub acute brain ischemia
induced vascular dementia. Eur J Pharmacol . 2014;723:80-90. http://dx.doi.org/10.1016/j.ejphar.2013.12.003
66. Hasselmo ME. The role of acetylcholine in
learning and memory. Curr Opin Neurobiol. 2006;16(6):710-715. https://dx.doi.org/10.1016%2Fj.conb.2006.09.002